

Leído para ud.: “¿Por qué una mosca no es un caballo?”. Evolución y ADN (3-3)

El Dogma cuestionado
Para que las proteínas funcionen bien no
solo deben tener la secuencia correcta de aminoácidos, sino también una
configuración espacial que las pliega de manera tal que se pueden
asociar entre ellas y con otras moléculas. Jacques Monod, premio nobel,
fue muy claro: “la información espacial necesaria para especifica la
estructura tridimensional de una proteína es inmensamente mayor que la
información contenida en la secuencia.”
Muchas proteínas fruto de la ingeniería
genética resultaron tener la forma incorrecta, y no funcionaron como se
suponía. El llamado mal de la “vaca loca” fue una de las consecuencias.
El agente fue una proteína “incorrecta”, proteínas priones.
La herencia genética es transmitida no
solo mediante el ADN. La continuidad de proteínas reveló una herencia en
paralelo que revoluciona el Dogma Central. La secuencia de aminoácidos
en las proteínas no es lo importante, sino su actitud en el espacio.
El misterio más grande de la biología ha
sido entender como un huevo se transforma en embrión. En el ser humano
esta recóndita morfogénesis ocurre en meras tres semanas cuando emerge
la organización espacial del organismo. Los huevos recién fertilizados
tienen una polaridad eléctrica. La corriente eléctrica fluye a través
del embrión, descargando acá y allá, provocando a las células a moverse,
plegarse y convulsionarse, creando nuevos patrones y más flujos y
contorsiones. La totalidad dinámica de estas fuerzas vectoriales se
conoce como campo morfogenético. En los núcleos de las
células inmersas en este minúsculo magma viviente, los genes se activan y
desactivan, proveyendo material para estos arcanos hornos. No son los genes quienes producen la forma naciente, sino la forma nesciente que selecciona y recluta los genes para su programa.
Pero lo más interesante de los campos
morfogenéticos es que se mantienen y transmiten a sí mismos
autónomamente, aparte de las dotes genéticas archivadas en el ADN.
Durante el desarrollo ambos sistemas se influyen mutuamente, el campo
despertando a los genes, y los genes proveyendo al campo con el material
necesario. La gran noticia biológica de fines del siglo XX fue
el reconocimiento de este segundo torrente hereditario, paralelo al
flujo del ADN.
Hasta ahora ha sido muy difícil descifrar
los campos morfogenéticos debido a su naturaleza tridimensional, su
turbulencia y su continua transformación. Pero es posible que sea en estos campos donde se guarden los secretos que distinguen una mosca de un caballo.
Las raíces secretas de las formas vivientes
El evolucionismo darwinista
se basa en la noción que las formas vivientes gradualmente divergen al
punto de convertirse en otras diferentes. Nadie jamás ha observado tal fenómeno.
El evolucionismo postula la necesidad de muchas “formas intermedias” en
el registro fósil, ramas que se habrían abierto durante millones de
años. A nivel molecular la teoría predice una serie de cambios adaptivos
(mutaciones) que supuestamente explicarían las diferencias morfológicas.
La biología molecular ha
certificado la unidad de los seres vivos, pero aparte de esto ha
decepcionado a las esperanzas evolucionistas. Los organismos difieren
ampliamente en su forma, sin embargo son celularmente similares y
bioquímicamente casi iguales. Es cierto que a mayor distancia entre dos
formas vivientes, mayores serán las diferencias moleculares –pero estas
diferencias tienen poco que ver con las formas. Ninguna molécula en la
almeja es más marina que su equivalente en el caballo. Ninguna molécula
del murciélago es más alada que la de la ballena.
Durante miles de millones de
años, se acuerdo a la escala de tiempo convencional del registro fósil,
la vida permaneció a nivel microscópico y unicelular. Luego en un
período geológico relativamente corto, casi como por encanto, la vida
animal que vino a poblar la tierra apareció en paralelo, en todos sus
tipos (i.e filos) morfológicos, incluyendo el nuestro (chordata). Esto es la llamada explosión cámbrica.
Debajo del cámbrico no hay ningún fósil que los pudo haber generado. El
mismo Darwin no pudo ofrecer ninguna explicación ante la carencia de
los presumidos ancestros pre-cámbricos “no puedo dar ninguna respuesta
satisfactoria.” Siglo y medio más tarde, y seguimos igual.
Esto demuestra que la vida
animal no surgió como en un delta, resultado de una transformación
gradual y diversificación de formas como postula el darwinismo –sino de
manera más bien abrupta, mediante “explosiones concertadas”. La
explosión de tipos (o filos) de vida no es un mero
insignificante e irresoluto detalle. Es exactamente lo opuesto a lo que
el gradualismo darwinista predijo sobre el origen de la formas de vida.
El filo de los vertebrados (estrictamente un subfilo) se divide en clases.
Todo intento de demostrar una derivación de las formas recientes a
partir de formas más antiguas, y de encontrar formas intermedias ha
resultado decepcionante. Los muy buscados “eslabones perdidos” siguen
extraviados.
El ambicioso proyecto
evolucionista de construir un Árbol de Vida genealógico, tanto a nivel
morfológico como molecular nunca se ha concretado. Por ejemplo los
estudios de la ya mencionada proteína citocromo c mostraron que no hay precedencia entre clases,
sino una derivación sincrónica (i.e. simultánea) a partir de un único
progenitor molecular. Los mamíferos, anfibios y peces resultaron ser
familiares distanciados, ninguno apareciendo en la escena antes que los
otros.
Uno de los hechos más
incómodos para los evolucionistas que salieron a la superficie gracias a
los estudios moleculares fue que todas las diferencias observables
entre proteínas homólogas entre varias clases son “neutrales” –no tienen
influencia en función o forma. La proteína citocromo c del pollo funciona perfectamente bien en la espinaca.
Volviendo a la evidencia ofrecida por los fósiles se ve que los mamíferos modernos (eutheria) son el ejemplo más impresionante de explosión vital. Más de 20 órdenes
de mamíferos emergieron casi al mismo tiempo, al comienzo del período
cenozoico (i.e. hace 65 millones, durante un periodo de solo unos 12
millones de años y 470 millones de años después que las formas de vida
animal aparecieran por primera vez). Los paleontólogos no han encontrado
gradaciones entre los órdenes de mamíferos.
Jamás se ha encontrado un
semi-murciélago. Nadie ha siquiera imaginado un buen ancestro común para
el fantástico bestiario de los órdenes mamíferos. El
árbol genealógico que muestra formas gradualmente variando y divergiendo
–propuesto por el darwinismo– simplemente no existe. Lo que si se
observa son arbustos sin raíces, apareciendo de la nada, con diferentes
flores que se parecen unos a otros, pero no se conocen entre sí.
¿Cómo podría ser la ancestral madre (o madres) común de todos estos
organismos? Tendría que ser una pequeña y elusiva larva, generalizada y
sin especializaciones, que contiene todo pero expresa casi nada, un
embrión ultra fecundo.
¿Larvas arcaicas madres desparramadas en
una vasta área flotando en su forma original hasta que un día son
convocadas a florecer? Como el bambú, que luego de muchos años verdes
florecen todos al mismo tiempo, en diferentes latitudes y altitudes, y
luego mueren. O como el avestruz, el ñandú, el emú, el casuarius y el
kiwi separados en las desconectadas sabanas de los continentes del sur.
Similares, pero desconocidos entre sí. ¿Quizás resultantes de una forma
arcaica que se desparramó en tiempos primordiales antes que los
continentes se separaran y luego geográficamente aislada en diferentes
lugares, se desarrolló autónomamente?
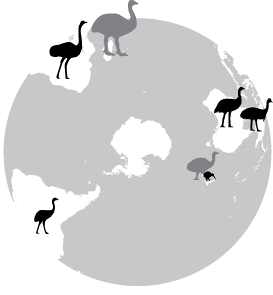 Avestruz
(África), ñandú (Sud América), emú (Australia), casuarius (Australia,
Nueva Guinea) y kiwi (Nueva Zelanda). El moa de Nueva Zelanda y el
pájaro elefante de Madagascar están ya extintos.
Avestruz
(África), ñandú (Sud América), emú (Australia), casuarius (Australia,
Nueva Guinea) y kiwi (Nueva Zelanda). El moa de Nueva Zelanda y el
pájaro elefante de Madagascar están ya extintos.Sermonti y Behe, coincidencias y divergencias
La biología ha develado en los últimos años tres grandes e independientes barreras contra el darwinismo[1] –o contra cualquier teoría donde el azar juega el rol principal.
La primera son las mutaciones aleatorias
mismas. El genoma codifica a muchos complejos sistemas moleculares. Las
mutaciones al azar en general rompen o dañan tales sistemas ya en
funcionamiento. A pesar de estos deterioros a veces los sistemas se
adaptan con la consiguiente propagación del error; algo que el trabajo
de Richard Lenski y su Long Term Evolution Project ha demostrado claramente.
El segundo obstáculo es la selección
natural misma. Como apuntó Darwin la selección natural trabaja sin
cesar, refinando un rasgo seleccionado para que cumpla cada vez mejor su
función. El problema es que a mayor especialización de un rasgo (o
herramienta), más difícil es usarlo para otro propósito, descartando la
ocurrencia de una mutación prohibitivamente improbable. Esto ha sido
demostrado por Joseph Thornton y su equipo, donde inclusive los más
modestos cambios en un sistema proteínico pre-existente son recibidos
con robustos e inesperados obstáculos evolutivos.
El tercer impedimento son las
complejidades irreducibles desarrolladas por Michael Behe. Ciertos
efectos selectivos necesitan más de una mutación antes que puedan
activarse. La probabilidad de múltiples mutaciones aleatorias decrece
exponencialmente con cada paso no-selectivo extra. Circunstancias
extraordinarias como alta tasa de mutación o enorme población en ciertos
virus y bacterias puede ayudar a saltar más de un paso al mismo tiempo,
pero son la excepción a la regla. En general unos pocos pasos no
selectivos ponen el asunto fuera del alcance de las mutaciones al azar.
Este impedimento es ubicuo a nivel molecular.
Las tres barreras son además independientes entre sí[2], y crean una poderosa sinergia contra la evolución aleatoria darwinista. En estos tres puntos Behe y Sermonti están de acuerdo.
En donde parecen divergir es en la
interpretación del supuesto ancestro común. Behe está más abierto a la
posible existencia de tal en el sentido que los darwinistas pretenden,
aunque descarta que tal evolución haya sido al azar. Sermonti es del
parecer que el ancestro no habría sido sino una versión original del ser
humano, al cual las mutaciones fueron gradualmente degenerando.
Conclusiones
Sermonti escribe con locuacidad
detallando no solo la inconsistencia evolucionista, la pobreza de la
ciencia reduccionista, y la arrogancia de los desdichados que con la
ingeniería genética se pensaron que eran dueños de la vida y de la
muerte; sino también contemplando atónito la majestad de la vida,
restituyéndole el misterio y el encanto que le son propios. En
particular subrayamos:
- La complejidad de los organismos no es proporcional al tamaño del genoma.
- La capacidad innata de la célula para armar proteínas aun cuando el gen que las codificas está incompleto.
- La incapacidad de las secuencias de ADN para explicar los pliegues y la tridimensionalidad de las proteínas, y demás formas y estructuras del organismo.
- Las inexplicadas homologías en especies no relacionadas, como el ojo del hombre y del pulpo. ¿Convergieron al azar hacia la misma estructura? Es lo que ahora postula la siempre cambiante teoría neo-darwiniana.
- La aparición súbita y consiguiente stasis de especies en el registro fósil, que destruye el deseado gradualismo darwinista; cuyos teóricos propusieron entonces el “equilibrio puntuado” para intentar explicar la cuadratura del círculo.
- Las cada vez más frecuentes “anomalías” filogenéticas las cuales indican que si existió un “ancestro común” entre monos y hombres, tendría que haber sido más similar a éste que aquel.
Mirar una vez más al organismo en su
conjunto, más allá de meras secuencia de ADN que no lo pueden explicar
totalmente, es donde Sermonti nos quiere conducir. Más importante aún,
cuestiona la muy promocionada –pero nunca probada– idea evolucionista
que lo simple e inferior deviene mediante procesos aleatorios complejo y
superior.
Recientemente se han propuesto nuevas
teorías evolutivas en la línea de Sermonti. Explican la evolución de la
vida en la tierra con mecanismos no-selectivos, como la PRM –Plasticidad-Relajación-Mutación
de Austin Hughes (2011). Aquí un organismo inicialmente con capacidad
“plástica” es capaz de sobrevivir en muchos medioambientes, pero cuando
se asienta en uno comienza a perder mediante mutaciones degradantes su
plasticidad primordial. Esto explicaría, entre otras cosas, las
características de “niño eterno” del ser humano, y su singular reloj
molecular. Por supuesto que no explica de dónde surgieron las funciones
primordiales y la vida misma. Pero apunta al hecho que muchas
grandes mentes de la biología están convergiendo a la idea que la
información necesaria para la vida estaba presente desde los mismos
comienzos, y que la vida se diversifica al perder gradualmente tal
información original. Esto es compatible con el concepto de creación divina, pero no con el evolucionismo darwinista.
Referencias
- Sermonti, G. 2005. Why is a Fly not a Horse? Dimenticare Darwin, Discovery Institute Press, 176 pp.
- Hughes, A. L., 2011 Evolution of adaptive phenotypic traits without positive Darwinian selection. Heredity (Edinb.) doi: 10.1038/hdy.2011.97.
Enrique de Zwart
Para Que no te la cuenten
Para descargar el artículo completo, hacer clic AQUÍ
[1] Aparte de la ya conocida carencia de fósiles que soporten la teoría darwinista.
[2] Para más detalles ver: Behe, M. The Edge of Evolution: Why Darwin’s Mechanism Is Self-Limiting
